Modelul unei situații „prădător-pradă”. Oscilații ale sistemului prădător-pradă (modelul Lotka-Voltaire) Modelarea prin simulare a sistemului „Predator-Prey”
Modelul lui Kolmogorov face o presupunere semnificativă: deoarece se presupune că aceasta înseamnă că există mecanisme în populația de pradă care le reglează numărul chiar și în absența prădătorilor.
Din păcate, o astfel de formulare a modelului nu permite să se răspundă la întrebarea în jurul căruia în ultima vreme Există multe dezbateri, despre care am menționat deja la începutul capitolului: cum poate o populație de prădători să exercite o influență de reglementare asupra unei populații de pradă, astfel încât întregul sistem să fie sustenabil? Prin urmare, vom reveni la modelul (2.1), în care mecanismele de autoreglare (de exemplu, reglarea prin competiție intraspecifică) sunt absente în populația de pradă (precum și în populația de prădători); prin urmare, singurul mecanism de reglare a numărului de specii incluse într-o comunitate este relația trofică dintre prădători și pradă.
Aici (deci, spre deosebire de modelul anterior, În mod firesc, soluțiile (2.1) depind de tipul specific de funcție trofică care, la rândul său, este determinată de natura prădării, adică de strategia trofică a prădătorului și de strategia defensivă a pradă comune tuturor acestor funcții (vezi Fig. I) sunt următoarele proprietăți:
Sistemul (2.1) are un punct staționar netrivial, ale cărui coordonate sunt determinate din ecuații
![]()
sub limitare naturală.
Mai există un punct staționar (0, 0), corespunzător echilibrului trivial. Este ușor de arătat că acest punct este o șa, iar separatoarele sunt axele de coordonate.
Ecuația caracteristică pentru un punct are forma
![]()
Evident, pentru modelul clasic Volterra.
Prin urmare, valoarea lui f poate fi considerată ca o măsură a abaterii modelului luat în considerare de la modelul Volterra.
![]()
un punct staționar este focalizarea, iar în sistem apar oscilații; când inegalitatea opusă este satisfăcută, există un nod și nu există oscilații în sistem. Stabilitatea acestei stări de echilibru este determinată de condiție
adică depinde în mod semnificativ de tipul funcției trofice a prădătorului.
Condiția (5.5) poate fi interpretată astfel: pentru stabilitatea echilibrului netrivial al sistemului prădător-pradă (și astfel pentru existența acestui sistem), este suficient ca în vecinătatea acestei stări proporția relativă de pradă consumată. de către prădător crește odată cu creșterea numărului de pradă. Într-adevăr, proporția de pradă (din numărul lor total) consumată de un prădător este descrisă de o funcție diferențiabilă, condiția pentru care să crească (derivat pozitiv) arată ca
![]()
Ultima condiție luată la punctul nu este altceva decât condiția (5.5) pentru stabilitatea echilibrului. Cu continuitate, trebuie îndeplinită și într-un anumit cartier al punctului. Astfel, dacă numărul victimelor din acest cartier, atunci
Fie că funcția trofică V are forma prezentată în Fig. 11, a (caracteristic nevertebratelor). Se poate demonstra că pentru toată lumea valorile finale(din moment ce convex in sus)
adică pentru orice valoare a numărului staționar de victime, inegalitatea (5.5) nu este satisfăcută.
Aceasta înseamnă că într-un sistem cu acest tip de funcție trofică nu există un echilibru stabil non-trivial. Sunt posibile mai multe rezultate: fie numărul de pradă, cât și de prădător crește la nesfârșit, fie (când traiectoria trece în apropierea uneia dintre axele de coordonate) din cauza motive aleatorii numărul de pradă sau numărul de prădători va deveni zero. Dacă prada moare, după ceva timp și prădătorul va muri, dar dacă prădătorul moare primul, atunci numărul de pradă va începe să crească exponențial. A treia opțiune - apariția unui ciclu limită stabil - este imposibilă, ceea ce este ușor de demonstrat.
De fapt, expresia

în cadranul pozitiv este întotdeauna pozitiv, cu excepția cazului în care are forma prezentată în Fig. 11, a. Apoi, conform criteriului Dulac, nu există traiectorii închise în această regiune și nu poate exista un ciclu limită stabil.
Deci, putem concluziona: dacă funcția trofică are forma prezentată în Fig. 11, iar atunci prădătorul nu poate fi un regulator care să asigure stabilitatea populației de pradă și, prin urmare, stabilitatea întregului sistem în ansamblu. Sistemul poate fi stabil doar dacă populația de pradă are propriile mecanisme interne de reglare, de ex. competiţie intraspecifică sau epizootii. Această opțiune de reglementare a fost deja discutată în §§ 3, 4.
S-a remarcat anterior că acest tip de funcție trofică este caracteristică pradătorilor de insecte, ale căror „victime” sunt, de asemenea, de obicei insecte. Pe de altă parte, observații ale dinamicii multora comunități naturale Tipurile „prădător-pradă”, care includ specii de insecte, arată că se caracterizează prin oscilații de amplitudine foarte mare și de tip foarte specific.
De obicei, după o creștere mai mult sau mai puțin treptată a numărului (care poate apărea fie monoton, fie sub formă de oscilații cu amplitudine crescândă), apare o scădere bruscă (Fig. 14), apoi imaginea se repetă. Aparent, această natură a dinamicii numărului de specii de insecte poate fi explicată prin instabilitatea acestui sistem la număr mic și mediu și acțiunea unor puternici regulatori intrapopulaționali ai numărului la număr mare.

Orez. 14. Dinamica populației psilidei australiane Cardiaspina albitextura hrănindu-se cu eucalipt. (Din articol: Clark L. R. The population dynamics of Cardiaspina albitextura.-Austr. J. Zool., 1964, 12, No. 3, p. 362-380.)
Dacă sistemul „prădător-pradă” include specii capabile de un comportament destul de complex (de exemplu, prădătorii sunt capabili să învețe sau prada sunt capabile să găsească adăpost), atunci un echilibru stabil non-trivial poate exista într-un astfel de sistem. Această afirmație este dovedită destul de simplu.
De fapt, funcția trofică ar trebui să aibă apoi forma prezentată în Fig. 11, c. Punctul de pe acest grafic este punctul de tangență al dreptei trasate de la originea graficului funcției trofice Evident, în acest punct funcția are un maxim. De asemenea, este ușor să arătăm că condiția (5.5) este îndeplinită pentru toți. În consecință, un echilibru netrivial în care numărul victimelor este mai mic va fi asimptotic stabil.
Cu toate acestea, nu putem spune nimic despre cât de mare este regiunea de stabilitate a acestui echilibru. De exemplu, dacă există un ciclu limită instabil, atunci această regiune trebuie să se afle în interiorul ciclului. Sau o altă opțiune: echilibrul netrivial (5.2) este instabil, dar există un ciclu limită stabil; în acest caz putem vorbi şi despre stabilitatea sistemului prădător-pradă. Deoarece expresia (5.7) atunci când alegeți o funcție trofică precum Fig. 11, în poate schimba semnul la schimbarea la , atunci criteriul Dulac nu funcționează aici și întrebarea existenței ciclurilor limită rămâne deschisă.

MODEL DE COMPUTER „PREDATOR-VICTIME”
Kazachkov Igor Alekseevici 1, Guseva Elena Nikolaevna 2
1 Statul Magnitogorsk universitate tehnică ei. G.I. Nosova, Institutul de Construcții, Arhitectură și Artă, student în anul V
2 Magnitogorsk State Technical University numit după. G.I. Nosova, Institutul de Energie și Sisteme Automatizate, Candidat la Științe Pedagogice, Conferențiar al Departamentului de Informatică de Afaceri și Tehnologii Informaționale
Adnotare
Acest articol este dedicat unei imagini de ansamblu asupra modelului computerizat „prădător-pradă”. Cercetările efectuate sugerează că modelarea mediului joacă un rol imens în cercetarea de mediu. Această problemă are mai multe fațete.
MODEL DE COMPUTER „PRADATOR-VICTIME”
Kazatchkov Igor Alekseevici 1, Guseva Elena Nikolaevna 2
1 Universitatea Tehnică de Stat Nosov Magnitogorsk, Institutul de Inginerie Civilă, Arhitectură și Arte, student al cursului al V-lea
2 Universitatea Tehnică de Stat Nosov Magnitogorsk, Institutul de Inginerie Energetică și Sisteme Automatizate, Doctor în Științe Pedagogice, Profesor asociat al Departamentului de Informatică de Afaceri și Tehnologii Informaționale
Abstract
Acest articol oferă o prezentare generală a modelului computerizat „prădător-victimă”. Studiul sugerează că simularea mediului joacă un rol imens în studiul mediului. Această problemă are mai multe fațete.
Modelarea ecologică este folosită pentru a studia mediul nostru. Modelele matematice sunt folosite în cazurile în care nu există mediu naturalși nu există obiecte naturale, ajută la prezicerea impactului diverși factori la obiectul studiat. Această metodă preia funcţiile de verificare, construire şi interpretare a rezultatelor obţinute. Pe baza unor astfel de forme, modelarea de mediu se ocupă de evaluarea schimbărilor din mediul din jurul nostru.
În momentul de față, astfel de forme sunt folosite pentru a studia mediul din jurul nostru, iar atunci când este necesar să studiem vreuna dintre domeniile acestuia, se folosește modelarea matematică. Acest model face posibilă prezicerea influenţei anumitor factori asupra obiectului de studiu. La un moment dat, tipul „prădător-pradă” a fost propus de oameni de știință precum: T. Malthus (Malthus 1798, Malthus 1905), Verhulst (Verhulst 1838), Pearl (Pearl 1927, 1930), precum și A. Lotka ( Lotka 1925, 1927) și V. Volterra (Volterra 1926) Aceste modele reproduc regimul oscilator periodic care apare ca urmare a interacțiunilor interspecifice din natură.
Una dintre principalele metode de cunoaștere este modelarea. Pe lângă faptul că poate prezice schimbările care apar în mediu, ajută și la găsirea cel mai bun mod rezolvarea problemei. Modelele matematice au fost folosite în ecologie de mult timp pentru a stabili modele și tendințe în dezvoltarea populațiilor și pentru a ajuta la evidențierea esenței observațiilor. Aspectul poate servi ca exemplu comportament, obiect.
La recrearea obiectelor în biologia matematică, se folosesc previziuni ale diferitelor sisteme, sunt prevăzute individualități speciale ale biosistemelor pentru: structura internă a individului, condițiile de susținere a vieții, constanța sistemelor ecologice, datorită cărora se păstrează activitatea vitală a sistemelor. .
Apariția modelării computerizate a avansat semnificativ granița capacităților de cercetare. A apărut posibilitatea implementării multilaterale a unor forme dificile care nu permit studiul analitic; cele mai noi direcții, precum și modelarea prin simulare.
Să luăm în considerare ce este un obiect de modelare. „Obiectul este un habitat închis în care două populații biologice interacționează: prădători și pradă. Are loc procesul de creștere, extincție și reproducere direct pe suprafața habitatului. Prada se hrănește cu resursele care sunt prezente în mediu, în timp ce prădătorii se hrănesc cu prada.
În acest caz, resursele nutriționale pot fi fie regenerabile, fie neregenerabile.
În 1931, Vito Volterra a derivat următoarele legi ale relației prădător-pradă.
Legea ciclului periodic - procesul de distrugere a prăzii de către un prădător duce adesea la fluctuații periodice ale mărimii populației ambelor specii, în funcție doar de rata de creștere a carnivorelor și ierbivorelor și de raportul inițial al numărului acestora. Legea conservării valorilor medii - numărul mediu al fiecărei specii este constant, indiferent de nivel de intrare
, cu condiția ca ratele specifice de creștere a populației, precum și eficiența prădării să fie constante.
Legea încălcării valorilor medii - atunci când ambele specii sunt reduse proporțional cu numărul lor, dimensiunea medie a populației de pradă crește, iar cea a prădătorilor scade. Modelul prădător-pradă este o relație specială între un prădător și prada acestuia, în urma căreia ambii beneficiază. Cei mai sănătoși și mai adaptați indivizi la condițiile de mediu supraviețuiesc, adică. toate acestea se întâmplă din cauza selecției naturale. Într-un mediu în care nu există oportunități de reproducere,
prădătorul va distruge mai devreme sau mai târziu populația prăzii, drept urmare ea însăși va dispărea.” Există multe organisme vii pe pământ care crește numărul rudelor în proporții enorme. Această abilitate se numește: potențialul biotic al unei specii, i.e. o creştere a numărului unei specii într-o anumită perioadă de timp. Fiecare specie are propriul său potențial biotic, de exemplu specii mari organismele într-un an pot crește de numai 1,1 ori, la rândul lor, organismele din specii mai mici, precum crustaceele etc. își pot crește aspectul de până la 1030 de ori, dar bacteriile sunt încă Mai mult. În oricare dintre aceste cazuri, populația va crește exponențial.
Creșterea exponențială a populației este o progresie geometrică a creșterii populației. Această capacitate poate fi observată în laborator în bacterii și drojdie. În condiții non-laboratoare, creșterea exponențială poate fi observată în exemplul lăcustelor sau la alte tipuri de insecte. O astfel de creștere a numărului de specii poate fi observată în acele locuri în care practic nu are dușmani și există hrană mai mult decât suficientă. În cele din urmă, expansiunea speciei, după ce numărul a crescut pentru o perioadă scurtă de timp, creșterea populației a început să scadă.
Să luăm în considerare un model computerizat de reproducere a mamiferelor folosind modelul Lotka-Volterra ca exemplu. Lasă Într-o anumită zonă trăiesc două tipuri de animale: căprioare și lupi. Modelul matematic al modificărilor populației în model Tavi-Volterra:
Numărul inițial de victime este xn, numărul de prădători este yn.
Parametrii modelului:
P1 – probabilitatea de a se întâlni cu un prădător,
P2 – coeficientul de creștere al prădătorilor în detrimentul prăzii,
d – rata mortalității prădătorilor,
a este coeficientul de creștere a numărului de victime.
În sarcina de antrenament, au fost stabilite următoarele valori: numărul de căprioare a fost 500, numărul de lup a fost 10, rata de creștere a căpriorului a fost 0,02, rata de creștere a lupilor a fost 0,1, probabilitatea de a întâlni un prădător a fost 0,0026, rata de creștere a prădătorilor în detrimentul prăzii a fost de 0,000056. Datele sunt calculate pentru 203 ani.
Explorăm influența coeficientul de creștere a victimelor pentru dezvoltarea a două populații, parametrii rămași vor rămâne neschimbați.În Schema 1 se observă o creștere a numărului de pradă și apoi, cu o oarecare întârziere, se observă o creștere a prădătorilor. Apoi prădătorii elimină victimele, numărul victimelor scade brusc și, în urma acestuia, numărul prădătorilor scade (Fig. 1).

Figura 1. Dimensiunea populației cu rate scăzute ale natalității în rândul victimelor
Să analizăm schimbarea modelului prin creșterea natalității victimei a=0,06. În Diagrama 2 vedem un proces oscilator ciclic care duce la o creștere a numărului ambelor populații în timp (Fig. 2).
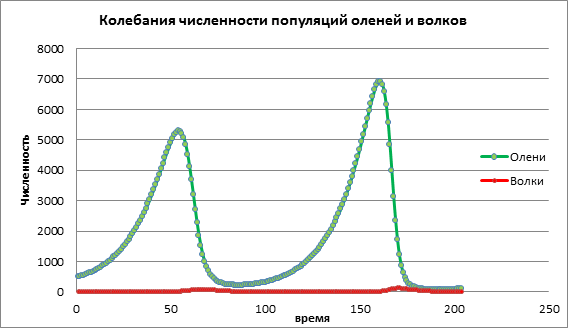
Figura 2. Dimensiunea populației la natalitatea medie a victimelor
Să luăm în considerare modul în care se va schimba dinamica populației cu o valoare ridicată a natalității victimei a=1,13. În fig. 3 se înregistrează o creștere bruscă a numărului ambelor populații, urmată de dispariția atât a prăzii, cât și a prădătorului. Acest lucru se întâmplă din cauza faptului că populația de pradă a crescut într-o asemenea măsură încât resursele au început să se epuizeze, ducând la dispariția prăzii. Extincția prădătorilor se produce din cauza faptului că numărul de pradă a scăzut, iar prădătorii au rămas fără resurse pentru a supraviețui.
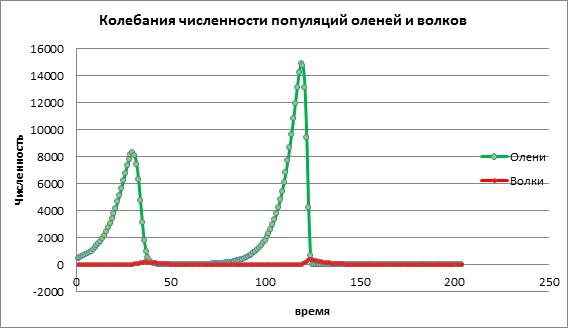
Figura 3. Dimensiunea populației cu rate ridicate ale natalității în rândul victimelor
Pe baza analizei datelor experimentale pe computer, putem concluziona că modelarea computerizată ne permite să prezicem mărimea populației și să studiem influența diverși factori asupra dinamicii populatiei. În exemplul de mai sus, am examinat modelul prădător-pradă, influența ratei natalității de pradă asupra numărului de căprioare și lup. O mică creștere a populației de pradă duce la o creștere mică a pradei, care după o anumită perioadă este distrusă de prădători. O creștere moderată a populației de pradă duce la creșterea dimensiunii ambelor populații. O creștere mare a populației de pradă duce mai întâi la o creștere rapidă a populației de pradă, aceasta afectând creșterea creșterii prădătorilor, dar apoi prădătorii care se înmulțesc distrug rapid populația de căprioare. Ca urmare, ambele specii dispar.
Adesea, membrii unei specii (populații) se hrănesc cu membrii unei alte specii.
Modelul Lotka-Volterra este un model al existenței reciproce a două populații de tip „prădător-pradă”.
Modelul prădător-pradă a fost dezvoltat pentru prima dată de A. Lotka în 1925, care l-a folosit pentru a descrie dinamica populațiilor biologice care interacționează. În 1926, independent de Lotka, modele similare (și mai complexe) au fost dezvoltate de către matematicianul italian V. Volterra, ale cărui cercetări profunde în domeniu probleme de mediu a pus bazele teorie matematică comunități biologice sau așa-numitele. ecologie matematică.
În formă matematică, sistemul de ecuații propus are forma:
unde x este numărul de pradă, y este numărul de prădători, t este timpul, α, β, γ, δ sunt coeficienți care reflectă interacțiunile dintre populații.
Enunțarea problemei
Luați în considerare un spațiu închis în care există două populații - ierbivore („pradă”) și prădători. Se crede că nu se importă sau exportă animale și că există suficientă hrană pentru ierbivore. Apoi, ecuația pentru modificarea numărului de victime (numai victime) va lua forma:
unde $α$ este rata natalității victimelor,
$x$ este dimensiunea populației de pradă,
$\frac(dx)(dt)$ este rata de creștere a populației de pradă.
Când prădătorii nu vânează, ei pot dispărea, ceea ce înseamnă că ecuația pentru numărul de prădători (numai prădătorii) devine:
Unde $γ$ este rata de pierdere a prădătorilor,
$y$ este dimensiunea populației de prădători,
$\frac(dy)(dt)$ este rata de creștere a populației de prădători.
Când prădătorii și prada se întâlnesc (frecvența întâlnirilor este direct proporțională cu produsul), prădătorii distrug victimele cu un coeficient, prădătorii bine hrăniți pot reproduce descendenți cu un coeficient; Astfel, sistemul de ecuații al modelului va lua forma:
Rezolvarea problemei
Să construim un model matematic al coexistenței a două populații biologice de tip „prădător-pradă”.
Lăsați două populații biologice să trăiască împreună într-un mediu izolat. Mediul este staționar și oferă cantități nelimitate din tot ceea ce este necesar pentru viață pentru una dintre specii - victime. O altă specie - un prădător - trăiește și ea în condiții staționare, dar se hrănește doar cu pradă. Pisicile, lupii, știucile, vulpile pot acționa ca prădători, iar găinile, iepurii de câmp, carasul și, respectiv, șoarecii pot acționa ca victime.
Pentru a fi specific, să considerăm pisicile ca prădători, iar găinile ca fiind victime.
Deci, puii și pisicile trăiesc într-un spațiu izolat - o curte a fermei. Mediul oferă găinilor hrană nelimitată, iar pisicile mănâncă numai găini. Să notăm prin
$x$ – numărul de pui,
$у$ – numărul de pisici.
În timp, numărul de găini și pisici se modifică, dar vom considera $x$ și $y$ ca fiind funcții continue ale timpului t. Să numim o pereche de numere $x, y)$ starea modelului.
Să aflăm cum se modifică starea modelului $(x, y).$
Să luăm în considerare $\frac(dx)(dt)$ – rata de modificare a numărului de găini.
Dacă nu există pisici, atunci numărul de găini crește și cu cât mai repede, cu atât sunt mai mulți pui. Vom presupune că dependența este liniară:
$\frac(dx)(dt) a_1 x$,
$a_1$ este un coeficient care depinde doar de condițiile de viață ale găinilor, de mortalitatea naturală a acestora și de rata natalității.
$\frac(dy)(dt)$ – rata de modificare a numărului de pisici (dacă nu există găini), depinde de numărul de pisici y.
Dacă nu există găini, atunci numărul pisicilor scade (nu au hrană) și se sting. Vom presupune că dependența este liniară:
$\frac(dy)(dt) - a_2 y$.
Într-un ecosistem, rata de modificare a numărului fiecărei specii va fi de asemenea considerată proporțională cu cantitatea acesteia, dar numai cu un coeficient în funcție de numărul de indivizi ai altei specii. Deci, la pui acest coeficient scade odată cu creșterea numărului de pisici, iar la pisici crește odată cu creșterea numărului de pui. De asemenea, vom presupune că dependența este liniară. Apoi obținem sistemul ecuatii diferentiale:
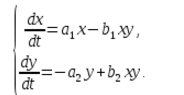
Acest sistem de ecuații se numește modelul Volterra-Lotka.
a1, a2, b1, b2 – coeficienți numerici, care se numesc parametri de model.
După cum puteți vedea, natura modificării stării modelului (x, y) este determinată de valorile parametrilor. Prin modificarea acestor parametri și prin rezolvarea sistemului de ecuații model, este posibil să se studieze modelele schimbărilor de stare sistem ecologic.
Folosind programul MATLAB, sistemul de ecuații Lotka-Volterra se rezolvă după cum urmează:
În fig. 1 prezintă soluția sistemului. În funcție de condițiile inițiale, soluțiile sunt diferite, ceea ce corespunde culori diferite traiectorii.
În fig. 2 prezintă aceleași soluții, dar ținând cont de axa timpului t (adică există o dependență de timp).
Dinamica populației este una dintre ramurile modelării matematice. Este interesant pentru că are aplicații specifice în biologie, ecologie, demografie și economie. Există mai multe modele de bază în această secțiune, dintre care unul, modelul „Predator-Prey”, este discutat în acest articol.
Primul exemplu de model în ecologia matematică a fost modelul propus de V. Volterra. El a fost primul care a considerat modelul relației dintre prădător și pradă.
Să luăm în considerare afirmația problemei. Să fie două tipuri de animale, dintre care unul îl devorează pe celălalt (prădători și pradă). În acest caz, se fac următoarele ipoteze: resursele alimentare ale prăzii nu sunt limitate și, prin urmare, în absența unui prădător, populația de pradă crește după o lege exponențială, în timp ce prădătorii, separați de victimele lor, mor treptat. de foame, tot după o lege exponenţială. Odată ce prădătorii și prada încep să trăiască în imediata apropiere unul de celălalt, schimbările în dimensiunea populației lor devin interdependente. În acest caz, evident, creșterea relativă a numărului de pradă va depinde de mărimea populației de prădători și invers.
În acest model, se presupune că toți prădătorii (și toate prada) sunt în aceleași condiții. În același timp, resursele alimentare ale victimelor sunt nelimitate, iar prădătorii se hrănesc exclusiv cu victime. Ambele populații trăiesc într-o zonă limitată și nu interacționează cu alte populații și nu există alți factori care ar putea afecta dimensiunea populației.
Modelul matematic „prădător-pradă” în sine constă dintr-o pereche de ecuații diferențiale care descriu dinamica populațiilor de prădători și pradă în cel mai simplu caz, când există o populație de prădători și una de pradă. Modelul este caracterizat de fluctuații ale mărimii ambelor populații, cu vârful la prădători ușor în spatele vârfului la pradă. Acest model poate fi găsit în multe lucrări despre dinamica populației sau modelare matematică. Acesta a fost acoperit și analizat pe scară largă folosind metode matematice. Cu toate acestea, formulele pot să nu ofere întotdeauna o idee evidentă a procesului care are loc.
Este interesant să aflăm cât de exact în acest model depinde dinamica populației de parametrii inițiali și cât de mult aceasta corespunde realității și bunul simț, și să-l vedeți grafic, fără a apela la calcule complexe. În acest scop, pe baza modelului Volterra, a fost creat un program în mediul Mathcad14.
Mai întâi, să verificăm modelul pentru conformitatea cu condițiile reale. Pentru a face acest lucru, să luăm în considerare cazurile degenerate când doar una dintre populații trăiește în condiții date. Teoretic, s-a demonstrat că, în absența prădătorilor, populația de pradă crește nelimitat în timp, iar populația de prădători în absența prăzii se stinge, ceea ce corespunde în general modelului și situației reale (cu formularea specificată a problemă).
Rezultatele obținute le reflectă pe cele teoretice: prădătorii se sting treptat (Fig. 1), iar numărul de pradă crește la nesfârșit (Fig. 2).
Fig. 1 Dependența numărului de prădători de timp în absența prăzii

Fig. 2 Dependența numărului de pradă în timp în absența prădătorilor
După cum se poate observa, în aceste cazuri sistemul corespunde modelului matematic.
Să luăm în considerare modul în care sistemul se comportă în funcție de diferiți parametri inițiali. Să fie două populații - lei și antilope - prădători și, respectiv, pradă, și se dau indicatorii inițiali. Apoi obținem următoarele rezultate (Fig. 3):
Tabelul 1. Coeficienții modului oscilator al sistemului

Fig.3 Sistem cu valorile parametrilor din tabelul 1
Să analizăm datele obținute pe baza graficelor. Odată cu creșterea inițială a populației de antilope, se observă o creștere a numărului de prădători. Rețineți că creșterea maximă a populației de prădători se observă mai târziu, în timpul scăderii populației de pradă, ceea ce este destul de consistent idei realeși modelul matematic. Într-adevăr, o creștere a numărului de antilope înseamnă o creștere a resurselor de hrană pentru lei, ceea ce presupune o creștere a numărului acestora. În plus, consumul activ de antilope de către lei duce la o scădere rapidă a numărului de pradă, ceea ce nu este surprinzător, având în vedere apetitul prădătorului, sau mai degrabă frecvența prădătorilor care mănâncă pradă. O scădere treptată a numărului de prădători duce la o situație în care populația de pradă se află în condiții favorabile creșterii. Apoi situația se repetă cu o anumită perioadă. Concluzionăm că aceste condiții nu sunt potrivite pentru dezvoltarea armonioasă a indivizilor, deoarece implică scăderi accentuate ale populației de pradă și creșteri puternice la ambele populații.
Să setăm acum numărul inițial al prădătorului egal cu 200 de indivizi menținând în același timp alți parametri (Fig. 4).
Tabelul 2. Coeficienții modului oscilator al sistemului

Fig.4 Sistem cu valorile parametrilor din tabelul 2
Acum sistemul oscilează mai natural. În aceste ipoteze, sistemul există destul de armonios, nu există creșteri și scăderi puternice ale numărului de numere în ambele populații. Concluzionăm că, cu acești parametri, ambele populații se dezvoltă suficient de uniform pentru a trăi împreună pe același teritoriu.
Să setăm numărul inițial de prădători la 100 de indivizi, numărul de pradă la 200, păstrând în același timp alți parametri (Fig. 5).
Tabelul 3. Coeficienții modului oscilator al sistemului

Fig.5 Sistem cu valorile parametrilor din Tabelul 3
ÎN în acest caz, situaţia este apropiată de prima situaţie luată în considerare. Rețineți că, odată cu creșterea reciprocă a populațiilor, trecerile de la o creștere la o scădere a populației de pradă au devenit mai lin, iar populația de prădători rămâne în absența prăzii la o valoare numerică mai mare. Concluzionăm că atunci când o populație este strâns legată de alta, interacțiunea lor are loc mai armonios dacă populațiile inițiale specifice sunt suficient de mari.
Să luăm în considerare modificarea altor parametri de sistem. Fie numerele inițiale să corespundă celui de-al doilea caz. Să creștem rata de reproducere a victimelor (Fig. 6).
Tabelul 4. Coeficienții modului oscilator al sistemului

Fig.6 Sistem cu valorile parametrilor din Tabelul 4
Să comparăm acest rezultat cu rezultatul obținut în al doilea caz. În acest caz, se observă o creștere mai rapidă a victimei. În acest caz, atât prădătorul, cât și prada se comportă ca în primul caz, ceea ce s-a explicat prin dimensiunea redusă a populației. Cu această interacțiune, ambele populații ajung la valori mult mai mari decât în al doilea caz.
Acum să creștem rata de creștere a prădătorilor (Fig. 7).
Tabelul 5. Coeficienții modului oscilator al sistemului

Fig.7 Sistem cu valorile parametrilor din Tabelul 5
Să comparăm rezultatele în mod similar. În acest caz caracteristici generale sistemul rămâne același, cu excepția modificării perioadei. După cum era de așteptat, perioada a devenit mai scurtă, ceea ce se explică prin scăderea rapidă a populației de prădători în absența prăzii.
Și, în sfârșit, să schimbăm coeficientul de interacțiune interspecifică. În primul rând, să creștem frecvența prădătorilor care mănâncă pradă:
Tabelul 6. Coeficienții modului oscilator al sistemului

Fig.8 Sistem cu valorile parametrilor din Tabelul 6
Deoarece prădătorul mănâncă prada mai des, numărul maxim al populației sale a crescut față de cel de-al doilea caz, iar diferența dintre maxim și valori minime mărimea populației. Perioada de oscilație a sistemului rămâne aceeași.
Și acum să reducem frecvența prădătorilor care mănâncă pradă:
Tabelul 7. Coeficienții modului oscilator al sistemului

Fig.9 Sistem cu valorile parametrilor din Tabelul 7
Acum prădătorul mănâncă prada mai rar, dimensiunea maximă a populației a scăzut față de cel de-al doilea caz, iar dimensiunea maximă a populației de pradă a crescut de 10 ori. Rezultă că, în aceste condiții, populația de pradă are o mai mare libertate în ceea ce privește reproducerea, deoarece prădătorul are nevoie de mai puțină masă pentru a obține suficientă. Diferența dintre dimensiunile maxime și minime ale populației a scăzut și ea.
Când se încearcă modelarea proceselor complexe din natură sau societate, într-un fel sau altul, se pune întrebarea cu privire la corectitudinea modelului. Desigur, la modelare, procesul este simplificat și unele detalii minore sunt neglijate. Pe de altă parte, există pericolul de a simplifica prea mult modelul, aruncând împreună cu cele neimportante caracteristici importante fenomene. Pentru a evita această situație, înainte de modelare este necesar să se studieze domeniul în care este utilizat acest model, să se examineze toate caracteristicile și parametrii acestuia și, cel mai important, să se evidențieze acele caracteristici care sunt cele mai semnificative. Procesul trebuie să aibă o descriere naturală, intuitivă de înțeles, coincizând în punctele principale cu modelul teoretic.
Modelul luat în considerare în această lucrare are o serie de dezavantaje semnificative. De exemplu, asumarea unor resurse nelimitate pentru victimă, absența unor factori terți care influențează mortalitatea ambelor specii etc. Toate aceste ipoteze nu reflectă situația reală. Cu toate acestea, în ciuda tuturor neajunsurilor, modelul a devenit larg răspândit în multe zone, chiar departe de ecologie. Acest lucru poate fi explicat prin faptul că sistemul „prădător-pradă” oferă o idee generală a interacțiunii speciilor. Interacțiunea cu mediu iar alți factori pot fi descriși de alte modele și analizați în combinație.
Relațiile prădător-pradă sunt o caracteristică esențială diverse tipuri activități de viață în care există o coliziune între două părți care interacționează. Acest model are loc nu numai în ecologie, ci și în economie, politică și alte domenii de activitate. De exemplu, unul dintre domeniile legate de economie este analiza pieței muncii, luând în considerare potențialii lucrători disponibili și locurile de muncă vacante. Acest subiect ar fi o continuare interesantă a lucrărilor la modelul prădător-pradă.
Agenția Federală pentru Educație
Instituție de învățământ de stat
studii profesionale superioare
„Universitatea Tehnică de Stat Izhevsk”
Facultatea de Matematică Aplicată
Departamentul „Modelarea matematică a proceselor și tehnologiilor”
Lucrări de curs
la disciplina „Ecuații diferențiale”
Subiect: „Cercetarea calitativă a modelului prădător-pradă”
Izhevsk 2010
INTRODUCERE
1. PARAMETRI ȘI ECUAȚIA DE BAZĂ ALE MODELULUI „PRĂDATOR-VICTICĂ”.
2.2 Modele Voltaire generalizate de tip „prădător-pradă”.
3. APLICAREA PRACTICĂ A MODELULUI „PRĂDATOR-VICTICĂ”.
CONCLUZIE
REFERINȚE
INTRODUCERE
În prezent, problemele de mediu sunt de o importanță capitală. Un pas important în rezolvarea acestor probleme este dezvoltarea modelelor matematice ale sistemelor ecologice.
Una dintre sarcinile principale ale ecologiei este scena modernă este studiul structurii și funcționării sistemelor naturale, căutare tipare generale. Mare influență Ecologia a fost influențată de matematică, care a contribuit la formarea ecologiei matematice, în special secțiuni precum teoria ecuațiilor diferențiale, teoria stabilității și teoria controlului optim.
Una dintre primele lucrări în domeniul ecologiei matematice a fost opera lui A.D. Lotki (1880 - 1949), care a fost primul care a descris interacțiunea diferitelor populații legate prin relații prădător-pradă. Mare contribuție V. Volterra (1860 - 1940), V.A a contribuit la studiul modelului prădător-pradă. Kostitsin (1883-1963) În prezent, ecuațiile care descriu interacțiunea populațiilor sunt numite ecuații Lotka-Volterra.
Ecuațiile Lotka-Volterra descriu dinamica valorilor medii - dimensiunea populației. În prezent, pe baza acestora, au fost construite modele mai generale de interacțiune a populației, descrise prin ecuații integro-diferențiale, și sunt studiate modele controlate prădător-pradă.
Unul dintre probleme importante ecologia matematică este problema sustenabilității ecosistemelor, managementului acestor sisteme. Managementul poate fi efectuat cu scopul de a transfera un sistem dintr-o stare stabilă în alta, în scopul utilizării sau refacerii acestuia.
1. PARAMETRII ȘI ECUAȚIA DE BAZĂ ALE MODELULUI PREDATOR-PRIMATE
Încercările de a modela matematic dinamica atât a populațiilor biologice individuale, cât și a comunităților, inclusiv a populațiilor care interacționează ale diferitelor specii, au fost întreprinse de mult timp. Unul dintre primele modele de creștere a populației izolate (2.1) a fost propus în 1798 de Thomas Malthus:
Acest model este specificat de următorii parametri:
N - dimensiunea populației;
Diferența dintre ratele natalității și cele ale mortalității.
Integrând această ecuație obținem:
![]() , (1.2)
, (1.2)
unde N(0) este mărimea populației la momentul t = 0. Evident, modelul Malthus la > 0 dă o creștere infinită a numărului, ceea ce nu se observă niciodată la populațiile naturale, unde resursele care asigură această creștere sunt întotdeauna limitate. Schimbările în numărul de populații de floră și faună nu pot fi descrise printr-o simplă lege malthusiană. proces de evolutie.
Descrierea matematică a acestor modele este tratată de ecologia matematică - știința relațiilor dintre organismele vegetale și animale și comunitățile pe care le formează între ele și cu mediul.
Cel mai serios studiu al modelelor de comunități biologice, inclusiv mai multe populații de specii diferite, a fost efectuat de matematicianul italian Vito Volterra:
 ,
,
unde este dimensiunea populației;
Ratele de creștere naturală (sau mortalitate) a unei populații; - coeficienţii de interacţiune interspecifică. În funcție de alegerea coeficienților, modelul descrie fie lupta speciilor pentru o resursă comună, fie o interacțiune prădător-pradă, atunci când o specie este hrană pentru alta. Dacă lucrările altor autori s-au concentrat pe construcția diferitelor modele, atunci V. Volterra a efectuat un studiu aprofundat al modelelor construite ale comunităților biologice. Odată cu cartea lui V. Volterra, conform multor oameni de știință, a început ecologia matematică modernă.
2. CERCETARE CALITATIVĂ A MODELULUI ELEMENTAL „PRĂDATOR-VICTICĂ”.
2.1 Model de interacțiune trofică în funcție de tipul „prădător-pradă”.
Să luăm în considerare modelul interacțiunii trofice de tip „prădător-pradă”, construit de V. Volterre. Să existe un sistem format din două specii, dintre care una o mănâncă pe cealaltă.
Luați în considerare cazul în care una dintre specii este un prădător și cealaltă este o pradă și vom presupune că prădătorul se hrănește numai cu pradă. Să acceptăm următoarea ipoteză simplă:
rata de creștere a victimelor;
Rata de creștere a prădătorilor;
Dimensiunea populației de pradă;
Dimensiunea populației de prădători;
Rata pradă de creștere naturală;
Rata consumului de pradă de către un prădător;
Rata mortalității unui prădător în absența prăzii;
Coeficientul de „prelucrare” a biomasei de pradă de către un prădător în propria sa biomasă.
Apoi, dinamica populației în sistemul prădător-pradă va fi descrisă printr-un sistem de ecuații diferențiale (2.1):
 (2.1)
(2.1)
unde toți coeficienții sunt pozitivi și constanți.
Modelul are o soluție de echilibru (2.2):
Conform modelului (2.1), ponderea prădătorilor în masa totală a animalelor este exprimată prin formula (2.3):
 (2.3)
(2.3)
O analiză a stabilității stării de echilibru în raport cu micile perturbări a arătat că punctul singular (2.2) este stabil „neutru” (de tip „centru”), adică orice abateri de la echilibru nu se sting, ci transferă sistem la un mod oscilator cu o amplitudine în funcție de magnitudinea perturbației. Traiectoriile sistemului pe planul de fază au forma unor curbe închise situate pe distante diferite din punctul de echilibru (Fig. 1).

Orez. 1 – Faza „portret” a sistemului clasic Volterra „prădător-pradă”
Împărțind prima ecuație a sistemului (2.1) la a doua, obținem ecuația diferențială (2.4) pentru curba pe planul de fază.
 (2.4)
(2.4)
Integrarea ecuația dată obținem:
![]() (2.5)
(2.5)
unde este constanta integrării, unde 
Este ușor de arătat că mișcarea unui punct de-a lungul planului de fază se va produce doar într-o singură direcție. Pentru a face acest lucru, este convenabil să înlocuiți funcțiile și prin mutarea originii coordonatelor pe plan într-un punct staționar (2.2) și apoi introducerea coordonatelor polare:
 (2.6)
(2.6)
În acest caz, înlocuind valorile sistemului (2.6) în sistemul (2.1), vom avea:
 (2.7)
(2.7)
Înmulțind prima ecuație cu și a doua cu și adunându-le, obținem:
După transformări algebrice similare obținem o ecuație pentru:
Cantitatea, după cum se poate vedea din (4.9), este întotdeauna mai mare decât zero. Astfel, nu se schimbă semnul, iar rotația este totul timpul trece Sens unic.
Integrând (2.9) găsim perioada:
Când sunt mici, ecuațiile (2.8) și (2.9) se transformă în ecuații ale unei elipse. Perioada de circulație în acest caz este egală cu:
 (2.11)
(2.11)
Pe baza periodicității soluțiilor ecuațiilor (2.1), putem obține unele consecințe. Pentru aceasta reprezentăm (2.1) sub forma:
 (2.12)
(2.12)
și să integreze pe parcursul perioadei:
 (2.13)
(2.13)
Deoarece substituțiile din și datorate periodicității dispar, mediile perioadei se dovedesc a fi egale cu stările staționare (2.14):
 (2.14)
(2.14)
Cele mai simple ecuații ale modelului „prădător-pradă” (2.1) au o serie de dezavantaje semnificative. Astfel, ei presupun resurse alimentare nelimitate pentru prada și creșterea nelimitată a prădătorului, ceea ce contrazice datele experimentale. În plus, după cum se poate observa din fig. 1, nici una dintre curbele de fază nu se distinge din punct de vedere al stabilității. În prezența unor influențe perturbatoare chiar și mici, traiectoria sistemului se va deplasa din ce în ce mai departe de poziția de echilibru, amplitudinea oscilațiilor va crește, iar sistemul se va prăbuși destul de repede.
În ciuda deficiențelor modelului (2.1), ideile despre natura fundamental oscilativă a dinamicii sistemului „ prădător-pradă» au devenit larg răspândite în ecologie. Interacțiunile prădător-pradă au fost folosite pentru a explica fenomene precum fluctuațiile numărului de animale răpitoare și pașnice din zonele de pescuit, fluctuațiile populațiilor de pești, insecte etc. De fapt, fluctuațiile numărului se pot datora și altor motive.
Să presupunem că în sistemul prădător-pradă există o distrugere artificială a indivizilor ambelor specii și să luăm în considerare întrebarea cum distrugerea indivizilor afectează valorile medii ale numărului lor dacă este efectuată proporțional cu acest număr, cu proporționalitate. coeficienţi şi, respectiv, pentru pradă şi prădător. Ținând cont de ipotezele făcute, rescriem sistemul de ecuații (2.1) sub forma:
 (2.15)
(2.15)
Să presupunem că, adică rata exterminării prăzii este mai mică decât rata creșterii sale naturale. În acest caz, se vor observa și fluctuații periodice ale numerelor. Să calculăm numerele medii:
 (2.16)
(2.16)
Astfel, dacă , atunci dimensiunea medie a populației de pradă crește, iar cea a prădătorului scade.
Să luăm în considerare cazul când coeficientul de exterminare a prăzii este mai mare decât coeficientul de creștere naturală a acesteia, adică. În acest caz ![]() pentru orice , și, prin urmare, soluția primei ecuații (2.15) este mărginită de sus de o funcție descrescătoare exponențial
pentru orice , și, prin urmare, soluția primei ecuații (2.15) este mărginită de sus de o funcție descrescătoare exponențial ![]() , adică la .
, adică la .
Pornind de la un anumit moment de timp t, la care , soluția celei de-a doua ecuații (2.15) începe și ea să scadă și tinde spre zero. Astfel, în cazul în care ambele specii dispar.
2.1 Modele Voltaire generalizate de tip „prădător-pradă”.
Primele modele ale lui V. Volterra, desigur, nu au putut reflecta toate aspectele interacțiunii în sistemul prădător-pradă, deoarece au fost mult simplificate în raport cu condițiile reale. De exemplu, dacă numărul unui prădător este zero, atunci din ecuațiile (1.4) rezultă că numărul de pradă crește la nesfârșit, ceea ce nu este adevărat. Valoarea acestor modele rezidă însă tocmai în faptul că ele au stat la baza pe care ecologia matematică a început să se dezvolte rapid.
Au apărut un număr mare de studii asupra diverselor modificări ale sistemului prădător-pradă, unde s-au construit modele mai generale care țin cont, într-o măsură sau alta, de situația reală din natură.
În 1936 A.N. Kolmogorov a propus utilizarea următorului sistem de ecuații pentru a descrie dinamica sistemului prădător-pradă:
 , (2.17)
, (2.17)
unde scade odată cu creșterea numărului de prădători și crește odată cu creșterea numărului de pradă.
Acest sistem de ecuații diferențiale, datorită generalității sale suficiente, face posibilă luarea în considerare a comportamentului real al populațiilor și, în același timp, realizarea analiza calitativa deciziile ei.
Mai târziu în lucrarea sa, Kolmogorov a explorat în detaliu un model mai puțin general:
 (2.18)
(2.18)
Diverse cazuri speciale ale sistemului de ecuații diferențiale (2.18) au fost studiate de mulți autori. Tabelul prezintă diverse cazuri speciale ale funcțiilor , , .
Tabelul 1 - Modele diverse comunități prădători-pradă
| Autorii | |||
| Volterra Lotka | |||
| Gause | |||
| Peaslow | |||
| Holing | |||
| Ivlev | |||
| Royama | |||
| Shimazu | |||
| mai |
modelare matematică pradă prădător
3. APLICAREA PRACTICĂ A MODELULUI PRĂDATOR-VICTICĂ
Să luăm în considerare un model matematic al coexistenței a două specii (populații) biologice de tip „prădător – pradă”, numit model Volterra – Lotka.
Lasă doi specii biologice trăiesc împreună într-un mediu izolat. Mediul este staționar și oferă cantități nelimitate din tot ceea ce este necesar pentru viață uneia dintre specii, pe care o vom numi victimă. O altă specie, un prădător, se află și ea în condiții staționare, dar se hrănește doar cu indivizii din prima specie. Aceștia ar putea fi carasul și știuca, iepuri de câmp și lupi, șoareci și vulpi, microbi și anticorpi, etc. Pentru claritate, le vom numi caras și știucă.
Sunt specificați următorii parametri inițiali:
De-a lungul timpului, numărul carasului și al știucii se modifică, dar întrucât în baltă sunt mulți pești, nu vom face distincția între 1020 caras și 1021 și de aceea îi vom considera și funcții continue ale timpului t. Vom numi o pereche de numere (,) starea modelului.
Este evident că natura schimbării stării (,) este determinată de valorile parametrilor. Prin modificarea parametrilor și rezolvarea sistemului de ecuații al modelului, este posibil să se studieze modelele de modificări ale stării sistemului ecologic în timp.
Într-un ecosistem, rata de modificare a numărului fiecărei specii va fi de asemenea considerată proporțională cu numărul acesteia, dar numai cu un coeficient care depinde de numărul de indivizi ai altei specii. Deci, la carasul acest coeficient scade odata cu cresterea numarului de stiuci, iar la stiuci creste odata cu cresterea numarului de caras. De asemenea, vom considera că această dependență este liniară. Apoi obținem un sistem de două ecuații diferențiale:

Acest sistem de ecuații se numește modelul Volterra-Lotka. Coeficienții numerici , , se numesc parametri de model. Este evident că natura schimbării stării (,) este determinată de valorile parametrilor. Prin modificarea acestor parametri și prin rezolvarea sistemului de ecuații model, este posibil să se studieze modelele de modificări ale stării sistemului ecologic.
Să integrăm ambele ecuații ale sistemului în raport cu t, care se va schimba de la momentul inițial de timp la , unde T este perioada în care au loc schimbări în ecosistem. Să fie în cazul nostru perioada de 1 an. Apoi sistemul ia următoarea formă:
 ;
;
 ;
;
Luând = și = și aducând termeni similari, obținem un sistem format din două ecuații:

Înlocuind datele inițiale în sistemul rezultat, obținem populația de știucă și caras din lac după un an: