Respiratory organs of arachnids. Type Arthropods
Respiratory system of spiders
Robert Gale Breen III
Southwestern College, Carlsbad, New Mexico, USA
Respiration, or the gas exchange of oxygen and carbon dioxide, in spiders is often not well understood even by experts. Many arachnologists, myself included, have studied various fields of entomology. Typically, courses in arthropod physiology center around insects. The most significant difference in the respiratory system of spiders and insects is that their blood or hemolymph plays no role in the respiration of insects, while in spiders it is a direct participant in the process.
Insect breath
The exchange of oxygen and carbon dioxide in insects is perfected largely due to the complex system of air tubes that make up the trachea and smaller tracheoles. Air tubes permeate the entire body in close contact with the internal tissues of the insect. For gas exchange between tissues and air tubes of an insect, hemolymph is not needed. This becomes clear in the behavior of certain insects, say, some species of grasshoppers. As the grasshopper moves, blood presumably circulates throughout the body as the heart stops. The blood pressure caused by the movement is enough for the hemolymph to perform its functions, which are to a greater extent the distribution of nutrients, water and the excretion of waste materials (a kind of equivalent to the kidneys of mammals). The heart starts beating again when the insect stops moving.
This is not the case with spiders, although it seems logical that spiders should proceed in this way, at least for those with tracheae.
Respiratory systems of spiders
Spiders have at least five different types of respiratory systems, depending on the taxonometric group and who you talk about it with:
1) The only pair of book lungs, like the haymakers Pholcidae;
2) Two pairs of book lungs - in the suborder Mesothelae and the vast majority of mygalomorph spiders (including tarantulas);
3) A pair of book lungs and a pair of tubular tracheae, as, for example, in weaver spiders, wolves, and most species of spiders.
4) A pair of tubular tracheae and a pair of sieve tracheas (or two pairs of tubular tracheae if you are one of those who are sure that the differences between tubular and sieve tracheas are not enough to distinguish them into separate species), as in a small family Caponiidae.
5) A single pair of sieve tracheas (or for some tubular tracheae), as in a small family Symphytognathidae.
Spider blood
Oxygen and carbon dioxide are transported along the hemolymph by the respiratory pigment protein, hemocyanin. Although hemocyanin is similar in chemical properties to vertebrate hemoglobin, unlike the latter, it contains two copper atoms, which gives the blood of spiders a bluish tint. Hemocyanin is not as effective at binding gases as hemoglobin, but its capabilities are quite enough for spiders.
As shown in the above depiction of a cephalothorax spider, the complex system of arteries leading to the legs and head region can be considered a predominantly closed system (according to Felix, 1996).
Spider trachea
Tracheal tubes penetrate the body (or parts of it, depending on the species) and end near the tissues. Yet this contact is not close enough for them to be able to supply oxygen and remove carbon dioxide from the body on their own, as happens in insects. Instead, the hemocyanin pigments have to pick up oxygen from the ends of the breathing tubes and pass it on, passing carbon dioxide back into the breathing tubes.
Tubular tracheae usually have one (rarely two) openings (called a spiracle or stigma), most of which open on the underside of the abdomen, next to the spinning appendages.
book lungs
The lung slits or book lung slits (in some species, the lung slits have various openings that can widen or narrow depending on oxygen demand) are located in front of the lower abdomen. The cavity behind the opening is stretched internally and accommodates many leaf-like air pockets of the book lung. The book lung is literally crammed with air pockets lined with an extremely thin cuticle that allows gas exchange through simple diffusion while blood flows through it. Tooth-like formations cover most of the surface of the book lungs on the side of the hemolymph flow to prevent collapse.
Digestive system of arachnids
How do spiders digest food?
» Arthropods » Arachnids » How do spiders digest food?
 Spiders kill or paralyze their prey by biting them and injecting venom through holes at the ends of their chelicerae. But chelicerae are unable to grind food into small pieces, and spiders have no teeth in their mouths. Therefore, spiders have adapted to eat liquid food. After killing the prey, the spider first injects its own digestive juices into it. In most animals, food is digested (broken down into simple substances) inside the body - in the stomach and intestines. This digestion is called internal. Spiders have external digestion: after a while, the tissues of the victim soften and turn into a nutrient solution that the spider absorbs, leaving only an empty skin.
Spiders kill or paralyze their prey by biting them and injecting venom through holes at the ends of their chelicerae. But chelicerae are unable to grind food into small pieces, and spiders have no teeth in their mouths. Therefore, spiders have adapted to eat liquid food. After killing the prey, the spider first injects its own digestive juices into it. In most animals, food is digested (broken down into simple substances) inside the body - in the stomach and intestines. This digestion is called internal. Spiders have external digestion: after a while, the tissues of the victim soften and turn into a nutrient solution that the spider absorbs, leaving only an empty skin.
spitter spiders, or hissing spiders (scytodes), catch prey by spraying it with a sticky liquid. Once on the victim, the liquid tightly sticks it to the substrate. The "glue" is produced by special glands in the back of the spider and is released into the air through the chelicerae. Kills prey with a bite.
Class Arachnids biology
Ability to match
Establish a correspondence between the signs and classes of animals for which these signs are characteristic: for each element of the first column, select the corresponding element from the second column.
Demonstration version of the Main state exam of the OGE 2017 - task 2017 - Task No. 25
FEATURES CLASSES
1) insects
2) arachnids
A) Some representatives in development have a pupal stage.
B) The vast majority of representatives are predators.
C) The body of an animal consists of a head, thorax and abdomen.
D) Animals can only eat liquid food.
D) Animals have four pairs of walking legs.
E) Simple and compound eyes can be located on the head.
Write in the table the selected numbers under the corresponding letters.
Solution:
Signs of Pa-at-about-different: pain-shin-stvo before-hundred-vi-te-lei - predator-no-ki; the body consists of the head-lo-chest and abdomen; able to consume only liquid food; have four pairs of walking legs; 8 simple eyes.
Signs on-se-ko-myh: there is a stage of ku-kol-ki (for some-one before-hundred-vi-te-lei), the body is co-hundred -it from the head, chest and abdomen, different types of mouths; have three pairs of walking legs; on the head, simple and complex eyes can diverge.
Answer: 121221
Respiratory, digestive, excretory system of spiders
Respiratory system
I think, after all that has been said, it will not surprise you that spiders also breathe differently.
Spiders in general can breathe with tracheae, book lungs, or both. The trachea is a system of thin tubes through which air reaches even distant parts of the spider's body. They are of little interest to us, since tarantulas and their closest relatives do not have tracheae.
But tarantulas have book lungs. There are 4 of them, and they resemble pockets on the underside of the opisthosoma, similar to the back pockets on jeans. Narrow openings are called pulmonary slits (also spiracles, stomata, stigmas). If you turn the tarantula over, then at least two of them (the rear pair) are visible. In well-fed individuals, the anterior pair is hidden by the basal segments of the last pair of legs. The lungs are clearly visible as white spots on the inside of the discarded opisthosoma exuvia. Inside the lungs are leaf-like folds of a thin membrane - lamellae ( lamellae, units lamella, also called leaflets or pages), which resemble the pages of a half-open book, hence the name. The hemolymph circulates within these folds, exchanging carbon dioxide for atmospheric oxygen, which separates the sheets from each other. The lamellas do not stick together with each other due to the many small spacers and racks. Book lungs are thought to be the result of the development of apodemes.
There has been much debate about the presence or absence of respiratory movements in tarantulas. Do they have active breathing with inhalation and exhalation, as we do? Supporters of this point of view point to the seemingly existing respiratory movements and muscles, which are closely associated with the lungs. Their opponents claim that tarantulas do not make breathing movements when observing them. For some reason, it so happened that the results of experiments conducted in this direction were contradictory or ambiguous. However, a series of experiments have recently been carried out and described (Paul et al. 1987), the results of which may put an end to the debate once and for all. It is shown that there are small fluctuations in the walls of the lungs, corresponding to the heartbeat and hemolymph pressure fluctuations.
But the additional volume of air attracted by these movements is so small that it does not play a significant role in gas exchange. Thus, the tarantula does not know the concept of inhalation and exhalation, relying entirely on diffusion.
Now that this riddle has been solved, we can still breathe a deep sigh of relief, although this is not given to tarantulas.
Digestive system
Spiders have no jaws. Instead, there are strong, strong chelicerae and fangs on them, and also rigid basal segments of pedipalps with spines and serrations. The mouth is located between the coxae of the pedipalps, directly above a small plate called the labium ( labium) or lower lip. The labium is a small outgrowth of the sternum (sternum). Above the mouth, between the bases of the chelicerae, there is another small plate, the labrum ( labrum) or upper lip. However, do not be misled: neither mobility nor function, these organs do not resemble human lips. It was simply more convenient for arachnologists of the past to give familiar names than to come up with something new, even more suitable.
Starting at the mouth, a narrow pharyngeal tube extends inward and upward, not very far. As soon as it reaches the anterior lower surface of the brain, it sharply bends horizontally and pierces it. (Remember the donut hole?) The horizontal section of the tube is called the esophagus.
The esophagus empties into a hollow muscular organ - the discharge stomach. The latter, with its elongated posterior end, connects to the real stomach, which lies between it and the brain. Finger-like protrusions extend from the real stomach to the bases of the legs - gastric (gastric) diverticula ( diverticula, units diverticulum).
The true stomach opens into a relatively straight-lying intestine, which enters the opisthosoma through the stalk.
Digestive and circulatory systems of arachnids
There, a bundle of filamentous organs, the Malpighian vessels, is connected to it. They perform the functions of the kidneys. Shortly before the gut opens into the anus, it forms a large protrusion, a blindly closed pouch called the stercoral pouch ( stercoral pocket). The anal opening is located directly above the arachnoid appendages. Tarantulas rely on the chelicerae, fangs, and coxae of the pedipalps for the difficult task of chewing their prey. Unlike them, other spiders pierce the integument of the victim and suck out the juices through a small hole.
Despite their large size, tarantulas consume only liquid food. Solid particles are filtered out by numerous hairs on the bases of the chelicerae and coxae of the pedipalps. Smaller particles, about a micron (0.001 mm) in size, are filtered out using the palatal plate, a special device in the throat. By comparison, most mammalian cells and most bacteria are larger than one micron. Spiders and most other arachnids do not like solid food.
While eating, tarantulas regurgitate digestive juices while simultaneously chewing their prey. The resulting slurry is diluted with the secretions of the coxal glands. As a result, partially digested liquid food is drawn into the mouth, then through the palatal plate into the pharynx and into the esophagus with the help of an injection stomach; in many ways it is similar to how we draw water through a straw, using the muscles of the cheeks and throat.
The pumping stomach is driven by powerful muscles, most of which are attached to the endosternitis and carapace. Through it, fluid from the esophagus flows back and down into the real stomach for further digestion and partial absorption. Finally, these processes are completed in the intestine. In the back of it, to what is left, waste products coming from the Malpighian vessels are added. All this accumulates in the stercoral pocket for some time. Periodically, excrement is excreted through the anus. Malpighian vessels are another example of parallel evolution. In spiders, they do not develop from the same embryonic structures as in insects. They were named after insects because they look almost the same, are located in almost the same place, and perform almost the same function. In short, these organs are analogous (similar but of different origin), not homologous (same origin and function).
Alternative names for parts of the digestive system are:
1.
rostrum (rostrum) instead of labrum;
2.
sucking stomach instead of delivery stomach;
3.
proximal midgut instead of true stomach;
4.
gastral cecum instead of gastral diverticulum;
5.
medial midgut instead of gut;
6.
cloacal chamber or cloaca instead of stercoral pocket and finally
7.
The posterior intestine is the short segment of the digestive tract between the sterocoral pocket and the anus.
Duplication of nomenclature occurs as a result of attempts to "fit" spiders to a measure taken from very different groups of arthropods, instead of developing a new one that best suits them.
Another aspect of spider digestion should also be discussed, namely the coxal glands. They belong to the digestive and excretory systems at the same time, so we talk about them at the intersection of these two topics.
Most arthropods possess coxal glands, which are direct homologs of the more primitive excretory organs, the nephridia, found in less advanced invertebrates. The tarantulas have them too. There are two pairs of them, and they are located on the backward side of the basal segments (coxae) of the 1st and 3rd pairs of legs, from which the name of these organs comes. For many years, arachnologists have struggled to figure out why they are needed. Many have been inclined to think that the coxal glands do not perform any function, being vestiges of more primitive nephridia that are no longer needed. The others weren't so sure. (Nefridii will be mentioned again on p. 46.)
Recently, Butt and Taylor (1991) determined that the coxal glands have a function. It seems that they secrete a saline solution to the mouth, seeping through the bends of the pleural membranes between the coxae and the sternum. This serves two purposes. Firstly, this ensures the liquid state of the food slurry that the tarantula drinks; this function is similar to that of our saliva. Secondly, this must be how the salt balance of the tarantula is maintained, since part of the salts is deposited in the dry residue of food. So, paradoxically, spiders salivate under the armpits!
The final well-chewed dry food residue mostly consists of inedible parts of the victim's body (i.e. exoskeleton), which the spider is not able to digest, as well as excess salts. Amateurs sometimes refer to this remnant as a riddle; professional arachnologists use the term food bolus.
In a large collection of tarantulas collected by the authors over many years (almost a thousand individuals at the moment), feeding is accompanied by a characteristic heavy sweet smell. It is not clear what causes this smell, digestive juices or overcooked food.
excretory system
One of the main problems of all animals is the timely removal of metabolic products before their concentration reaches a dangerous level. Digestible substances consist mainly of carbon, hydrogen, oxygen and nitrogen with traces of other elements. During metabolism, carbon is converted to carbon dioxide and excreted through the lungs or gills. Hydrogen becomes water, which is no different from the water that enters the body with food or drink. Oxygen can be incorporated into various organic compounds or removed as part of carbon dioxide.
The hardest part is nitrogen.
Together with hydrogen, it gives ammonia, a very toxic compound. Aquatic animals can get rid of nitrogen in the form of ammonia or other soluble substances by simply allowing them to dissolve in the surrounding water. They usually have plenty of water and little energy is spent on excretion.
Land animals are not so lucky. If nothing is done, the concentration of nitrogen compounds quickly increases to lethal. Several methods have been devised to avoid poisoning. The first is to convert nitrogen into a less toxic form than ammonia. If this product is less soluble, then even more of it can be accumulated if concentrated. And if there is still an opportunity to isolate the concentrate from the internal environment of the body, then it becomes much safer. Finally, the ideal end product should be easy to hatch with a minimum of water, salt and energy consumption.
Arachnids in general, and spiders in particular, have developed a technology that combines all of these approaches. And they did it again in their own way.
First, it is necessary to develop a relatively safe substance. The main excreted product in spiders is guanine, other nitrogen-containing wastes (adenine, hypoxanthine, uric acid) are excreted in small quantities. In this, arachnids stand in stark contrast to the rest of the animal kingdom, which never excrete guanine as waste (Anderson 1966; Rao and Gopalakrishnareddy 1962). Although they also produce it, rest assured. In cats and deer, for example, guanine is the main substance that provides the reflective properties of the retina. But, unlike spiders, cats and deer do not excrete it as waste. Since guanine is insoluble, it is completely harmless to the spider.
Again, because it is insoluble, it can deposit as a solid and accumulate more efficiently. Compared to urea, for example, it takes up much less space and needs to be disposed of less frequently. Then, since it is a solid, you can store it in safe places. Some intestinal cells (so-called guanocytes) are able to accumulate quite large amounts of guanine. Although they do not remove guanine from the body, they effectively neutralize it, allowing the body to function without worrying about the energy and material costs of excretion.
And finally, by concentrating waste products to a solid state, the spider can get rid of them with little loss of water, salts and energy. B about Most of the guanine secreted by the Malpighian vessels accumulates in the stercoral pocket and is ejected from there along with the remnants of undigested food. Thus, arachnids (and spiders among them) use all 4 approaches to avoid nitrogen poisoning, and they do it extremely effectively.
An interesting consequence of all of the above is that spiders do not have kidneys, they do not produce urine, which means they are not familiar with the concept urinate, at least in the sense in which we usually use it. In that case, what do they do?
reproductive system
The sexual life of tarantulas is truly stunning, but it will be discussed a little later. Here, we restrict ourselves to a simple description of the mechanism.
The gonads of spiders - the ovaries in females and the testes in males - are located inside the opisthosome. Single genital opening (gonopore, gonopore) is located on the ventral surface of the opisthosome and is located along a groove called epigastric groove, which runs in the transverse direction, connecting the upper lungs. This is the posterior edge of the epiginal plate. In early literature, the epigastric sulcus is sometimes referred to as the generative fold. In the female, two ovaries are connected to a single oviduct, which opens with a gonopore. Directly inside the gonopore are two "pockets", which are called spermatheca or spermatheca ( spermathecae, units spermatheca). During copulation (mating), the male deposits the sperm into the spermatheca, where the spermatozoa remain alive until they need to fertilize the eggs, weeks or months later.
In the male, the paired testicles are spirally twisted tubes that open into a common duct. The duct, in turn, opens into the outside world again by the gonopore. Next to the gonopore are the epiandral glands; it is believed that they either contribute to the formation of seminal fluid, or develop a special thread for weaving the sperm web (Melchers 1964).
The male spider does not have a penis or any homologous organ. Its copulatory appendages are the secondary sex organs at the ends of the pedipalps. In adult males, the terminal segment of the pedipalp (pretarsus and claw) transforms from a simple construct seen in immature males to a complex, highly specialized organ for introducing sperm into the female genital tract. This segment resembles an exotic bottle, bulbous, with an elaborately curved and twisted neck. The body of the bottle is called the bulb ( bulb) or a reservoir, and the neck is an embolus ( embolus, pl. emboli). The foot, meanwhile, shortens and thickens. The embolus and bulb are attached to it with a flexible articulation that allows them to move freely in different planes. The modified foot is often called a cymbium ( cymbium, pl. cymbia). The cymbium is connected to the tibia by another elastic joint.
Berce bears a special groove (alveolus, alveolus), the shape of which corresponds to the shape of the embolus and bulb. Thanks to the mobility of the cymbium, the spider can tuck them into this groove when they are not needed. But when the embolus and bulb are filled with sperm and ready to be injected into the female genital tract, they are completely open and turned at the right angle with respect to the pedipalp.
This class includes arthropods adapted to living on land, breathing through the lungs and trachea. The class unites the detachments of spiders, ticks, scorpions, haymakers.
a brief description of
|
body structure |
Body consists of cephalothorax and abdomen |
|
body integuments |
Body covered with chitinous cuticle |
|
limbs |
On the cephalothorax - 6 pairs of limbs: 2 pairs of jaws, 4 pairs of walking legs. No antennas or antennules |
|
body cavity |
Mixed cavity of the body, in which the internal organs are located |
|
Digestive system |
Anterior gut. Pharynx. Midgut. Hind gut. Liver. Spiders have partially external digestion |
|
Respiratory system |
Lungs or trachea |
|
Circulatory system |
The heart is in the form of a tube with lateral slit-like processes - ostia. The circulatory system is not closed. Hemolymph contains the respiratory pigment hemocyanin |
|
excretorysystem |
Malpighian vessels |
|
Nervous system |
Consists of the brain - supraglottic node, peripharyngeal ring, abdominal nerve chain |
|
sense organs |
Sensitive hairs, which are especially numerous on the pedipalps. The organs of vision are represented by simple eyes from 2 to 12 |
|
Reproductive system and development |
Arachnids have separate sexes. Fertilization is internal. Pronounced sexual dimorphism |
general characteristics
Structure and integument. For arachnids, a characteristic feature is the tendency to merge the segments of the body that form the cephalothorax and abdomen. Scorpions have a fused cephalothorax and a segmented abdomen. In spiders, both the cephalothorax and the abdomen are continuous undivided sections of the body, between which there is a short stalk connecting these two sections. The maximum degree of fusion of body segments is observed in ticks, which have lost even the division of the body into the cephalothorax and abdomen. The body of ticks becomes whole without borders between segments and without constrictions.
The integuments of arachnids consist of the cuticle, hypodermis, and basement membrane. The outer layer of the cuticle is a lipoprotein layer. This layer protects very well from loss of moisture during evaporation. In this regard, arachnids were able to become a real terrestrial group and settle in the most arid regions of the earth. The composition of the cuticle also includes proteins, tanned with phenols and encrusting chitin, which gives the cuticle strength. Derivatives of the hypodermis are spider and poisonous glands.
Limbs. Head limbs, except for two pairs of jaws, are absent in arachnids. The jaws, as a rule, are attributed to the limbs of the cephalothorax. The cephalothorax of arachnids bears 6 pairs of limbs, which is a distinctive feature of this class. The two front pairs are fitted
to capture and grind food - chelicerae and pedipalps (Fig. 1). Chelicerae, which look like short claws, are located in front of the mouth. In spiders, chelicerae end in a claw, near the top of which is the opening of the poisonous gland. The second pair is pedipalps, on the main segment they have a chewing outgrowth, with the help of which food is crushed and kneaded. In some species, pedipalps turn into powerful claws (for example, in scorpions) or look like walking legs, and in some forms of spiders, a copulatory organ may be located at the end of the pedipalps. The remaining 4 pairs of limbs of the cephalothorax perform the function of movement - these are walking legs. During embryonic development, a large number of limbs are laid on the abdomen, but in adult chelicerates, the abdomen is devoid of typical limbs. If the abdominal limbs persist into adulthood, they are usually modified into genital opercula, tactile appendages (scorpions), lung sacs, or arachnoid warts.

Rice. one. The mouth organs of the cross spider: 1 - the terminal claw-like segment of the chelicera; 2 - basal segment of helicerae; 3 - pedipalp; 4 - chewing outgrowth of the main segment of the pedi-palp; 5 - main segment of the walking leg
The digestive system (Fig. 2) has features associated with a peculiar way of eating arachnids - extraintestinal, or external, digestion. Arachnids cannot take solid food in chunks. Digestive enzymes are injected into the victim's body and turn its contents into a liquid slurry that is absorbed. In this regard, the pharynx has strong muscles and serves as a kind of pump that draws in semi-liquid food. The midgut of most arachnids has lateral blind protrusions to increase the absorptive surface. In the abdomen, the ducts of the paired liver open into the intestine. The liver performs not only digestive functions, releasing digestive enzymes, but also an absorption function. Intracellular digestion takes place in the liver cells. The hindgut ends at the anus.
The respiratory system of arachnids is represented by lung sacs and tracheae. At the same time, some species have only lung sacs (scorpions, primitive spiders). In others, the respiratory organs are represented only by tracheae.

2. Spider organization scheme: 1 - eyes; 2 - poisonous gland; 3 - chelicera; 4 - brain; 5 - mouth; 6 - subpharyngeal nerve node; 7 - glandular outgrowth of the intestine; 8 - bases of walking legs; 9 - lung; 10 - pulmonary opening - spiracle; 11 - oviduct; 12 - ovary; 13 - spider glands; 14 - arachnoid warts; 15 - anus; 16 - Malpighian vessels; 17 - os-ti; 18 - ducts of the liver; 19 - heart; 20 - pharynx connected with the body wall by muscles
(salpugs, haymakers, some ticks). In spiders, two types of respiratory organs occur simultaneously. There are four-lung spiders that have 2 pairs of lung sacs and no tracheae; bipulmonary spiders - one pair of lung sacs and a pair of tracheal bundles and lungless spiders - only tracheas. Some small spiders and some mites have no respiratory organs and respiration is carried out through the thin integuments of the body.
Circulatory system, as in all arthropods, open. Hemolymph contains the respiratory enzyme hemocyanin.

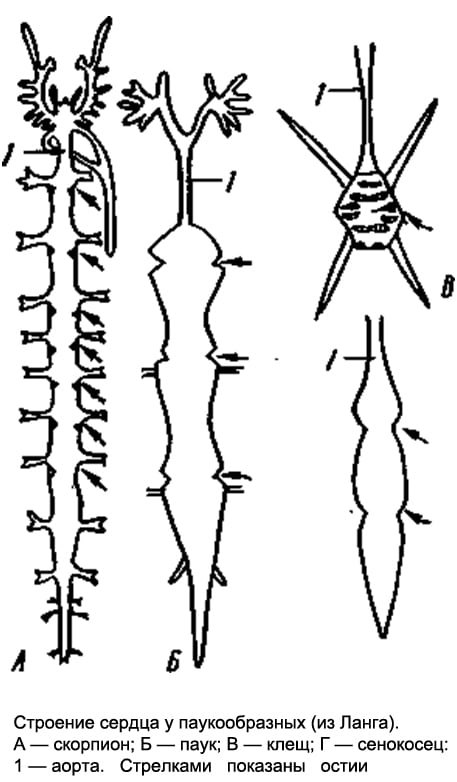
Rice. 3. The structure of the heart in arachnids. A - scorpion; B - spider; B - tick; G - haymaker: 1 - aorta (arrows show ostia)
The structure of the heart depends on the degree of segmentation - the more segments, the more ostia (Fig. 3). In ticks that lack segmentation, the heart may completely disappear.
excretory system in adult arachnids, it is represented by a pair of branching Malpighian vessels that open at the border of the middle and hind intestines into the digestive system.
Nervous system arachnids, like circulatory, depends on the segmentation of the body. The least concentrated nerve chain in scorpions. In arachnids, unlike crustaceans and insects, the brain consists of two sections - anterior and posterior, the middle section of the brain is absent, since arachnids do not have head limbs, antennules or antennae, which this section should control. There is a large ganglionic mass in the cephalothorax and ganglia of the abdominal chain. With a decrease in segmentation, the ventral chain disappears. So, in spiders, the entire abdominal chain merges into the cephalothoracic ganglion. And in harvestmen and ticks, the brain and cephalothoracic ganglion form a continuous ganglionic ring around the esophagus.
sense organs mainly represented by special hairs that are located on the pedipalps, legs and surface of the body and respond to air vibrations. On the pedipalps there are also sensory organs that perceive mechanical and tactile stimuli. The organs of vision are represented by simple eyes. The number of eyes can be 12, 8, 6, rarely 2.
Development. Most arachnids lay eggs, but live births have also been observed. Development is direct, but ticks have metamorphosis.
A.G. Lebedev "Preparing for the exam in biology"
excretory system. The excretory system is represented by the Malpighian vessels, which are a neoplasm in Arachnoidea, and the coxal glands, which correspond to coelomoducts. Malpighian vessels - a pair of branching, blindly closed at the ends of the tubes, open at the border of the middle and hind intestines.
They are of endodermal origin, that is, they belong to the middle intestine. Grains of guanine, the main excretory product of arachnids, accumulate in the epithelium and lumen of the Malpighian vessels. The coxal glands are formed by the sac-like part of mesodermal origin, the convoluted duct (labyrinth), the reservoir, and the external excretory duct. They are available in one or two pairs, open at the bases of the legs and rarely function in adult forms.
reproductive system. Arachnids have separate sexes. The sex glands are located in the abdomen and in the initial state of the pair. In some cases, there is a fusion of the right and left gonads. So, in a male scorpion, the testes are paired and each consists of two tubes connected by jumpers; in female scorpions, the ovary is one and consists of three tubes, of which the middle tube is obviously the result of the fusion of two medial tubes, similar to those of the male. In many spiders, harvestmen, and ticks, the paired gonads grow together at the ends into a ring. Paired oviducts and seminal ducts open with an unpaired genital opening, always on the second segment of the abdomen. The structure of the excretory part of the reproductive system and the copulatory adaptations of males are very diverse. Females usually have an extension of the oviducts - the uterus and seminal receptacles. In males, the copulatory organs are either associated with the genital opening, orserve as pedipalps (spiders) or chelicerae (some mites). In some cases, spermatophoric fertilization is carried out with the help of sperm packets.
Development. Most arachnids lay eggs, but there are also viviparous forms (scorpions, some ticks, etc.). Eggs are richyolk, due to which fragmentation is partial, superficial, all segments of the body and limbs are formed in embryonic development, and a small full-segment individual, similar to an adult, hatches from the egg. Post-embryonic development is direct, accompanied mainly by growth. Only in ticks, due to the small size of the eggs, a six-legged larva hatches and metamorphosis takes place. The study of the embryos of primitive arachnids allows us to better understand the structure of adults. So, in the embryo of scorpions, abdominal limbs are laid on all segments of the mesosome, from which the first pair then disappears, the second turns into genital covers, the third into ridge-shaped organs, and the remaining four pairs into lungs.
Respiratory system. The respiratory organs of the cross are a pair of leaf-folded lungs and tubular tracheae. The lungs are located at the base of the abdomen on the sides of the genital opening, where there are two transverse slits - stigmas of the lungs.
The stigma leads to the lung cavity, on the wall of which there are a number of flat pockets that diverge in a fan-like fashion. The pockets are connected with jumpers and do not fall off, so that air freely penetrates between them. Blood circulates in the cavities of the pockets, the exchange of gases occurs through their thin cuticular walls.
The tracheal system consists of two non-branching tubes, which are directed forward from a common pocket, which opens with an inconspicuous transverse slit in front of the arachnoid warts.
excretory system. There are two types of excretory organs: Malpighian vessels and coxal glands. In addition, the excretory function is performed by special cells (nephrocytes and guanocytes) lying in the body cavity. The Malpighian vessels are represented by four branching, blindly closed tubes at the ends, which flow into the rectal bladder along its sides at the border of the middle and posterior intestines. Malpighian vessels are lined with squamous epithelium, in the cells of which grains of guanine, the main excretion product, are formed. The coxal glands, which in arachnids are the remains of the coelomoduct system, are located at the base of the first pair of legs. In an adult spider, they do not function.
poison glands. Poisonous glands are located in the anterior part of the cephalothorax at the base of the chelicerae. This is a pair of rather large cylindrical glands that enter the cavity of the main segments of the chelicerae. The outer lining of the gland is formed by a spirally curled ribbon-like muscle, during the contraction of which the poison is poured out through a thin duct that opens at the end of the claw-like segment of the chelicerae.
Spinning apparatus. The spinning apparatus is represented by three pairs of spider warts and spider glands. At rest, spider warts, together with the anal tubercle, form a common closed group. At the tops of the warts there are numerous arachnoid tubes through which a secret is secreted - a web that hardens when it comes into contact with air. Spider glands fill the lower part of the female's abdominal cavity.
Their structure and size are not the same; distinguish tubular, ampulloidal, dendritic and pear-shaped glands. The latter are especially numerous and connected in bundles according to the number of warts (Plate X). The role of various glands and warts is different, the tubular glands secrete a web for the egg cocoon, the ampulloidal glands for building a network, the pear-shaped glands for braiding prey; arboreal secrete a sticky secret that covers the network.
More interesting articles
Respiratory system of spiders
Robert Gale Breen III
Southwestern College, Carlsbad, New Mexico, USA
Respiration, or the gas exchange of oxygen and carbon dioxide, in spiders is often not well understood even by experts. Many arachnologists, myself included, have studied various fields of entomology. Typically, courses in arthropod physiology center around insects. The most significant difference in the respiratory system of spiders and insects is that their blood or hemolymph plays no role in the respiration of insects, while in spiders it is a direct participant in the process.
Insect breath
The exchange of oxygen and carbon dioxide in insects is perfected largely due to the complex system of air tubes that make up the trachea and smaller tracheoles. Air tubes permeate the entire body in close contact with the internal tissues of the insect. For gas exchange between tissues and air tubes of an insect, hemolymph is not needed. This becomes clear in the behavior of certain insects, say, some species of grasshoppers. As the grasshopper moves, blood presumably circulates throughout the body as the heart stops. The blood pressure caused by the movement is enough for the hemolymph to perform its functions, which are to a greater extent the distribution of nutrients, water and the excretion of waste materials (a kind of equivalent to the kidneys of mammals). The heart starts beating again when the insect stops moving.
This is not the case with spiders, although it seems logical that spiders should proceed in this way, at least for those with tracheae.
Respiratory systems of spiders
Spiders have at least five different types of respiratory systems, depending on the taxonometric group and who you talk about it with:
1) The only pair of book lungs, like the haymakers Pholcidae;
2) Two pairs of book lungs - in the suborder Mesothelae and the vast majority of mygalomorph spiders (including tarantulas);
3) A pair of book lungs and a pair of tubular tracheae, as, for example, in weaver spiders, wolves, and most species of spiders.
4) A pair of tubular tracheae and a pair of sieve tracheas (or two pairs of tubular tracheae if you are one of those who are sure that the differences between tubular and sieve tracheas are not enough to distinguish them into separate species), as in a small family Caponiidae.
5) A single pair of sieve tracheas (or for some tubular tracheae), as in a small family Symphytognathidae.
Spider blood
Oxygen and carbon dioxide are carried along the hemolymph by the respiratory pigment protein, hemocyanin. Although hemocyanin is similar in chemical properties to vertebrate hemoglobin, unlike the latter, it contains two copper atoms, which gives the blood of spiders a bluish tint. Hemocyanin is not as effective at binding gases as hemoglobin, but its capabilities are quite enough for spiders.
As shown in the above depiction of a cephalothorax spider, the complex system of arteries leading to the legs and head region can be considered a predominantly closed system (according to Felix, 1996).
Spider trachea
Tracheal tubes penetrate the body (or parts of it, depending on the species) and end near the tissues. Yet this contact is not close enough for them to be able to supply oxygen and remove carbon dioxide from the body on their own, as happens in insects. Instead, the hemocyanin pigments have to pick up oxygen from the ends of the breathing tubes and pass it on, passing carbon dioxide back into the breathing tubes. Tubular tracheae usually have one (rarely two) openings (called a spiracle or stigma), most of which open on the underside of the abdomen, next to the spinning appendages.
book lungs
The lung slits or book lung slits (in some species, the lung slits have various openings that can widen or narrow depending on oxygen demand) are located in front of the lower abdomen. The book lung is literally crammed with air pockets lined with an extremely thin cuticle that allows gas exchange through simple diffusion while blood flows through it. Tooth-like formations cover most of the surface of the book lungs on the side of the hemolymph flow to prevent collapse.
Breath of tarantulas
Since tarantulas are large and easier to study, many physiologists, when considering the mechanism of respiration of spiders, dwell on them. The geographical habitat of the studied species is rarely specified, it can be assumed that most of them come from the United States. Almost universally, the taxonomy of tarantulas is not taken into account. Only rarely do physiologists engage a competent spider taxonomist. More often, they believe anyone who says they can identify test species. This disregard for systematics is evident even among the most famous physiologists, including R.F. Felix, author of the only widely circulated, but, alas, not the most accurate book on spider biology.
Book lung composed of leaf-like intermittent air pockets with venous hemolymph flowing in one direction between the pockets. The layer of cells that isolate the air pockets from the hemolymph is so thin that gas exchange by diffusion becomes possible (according to Felix, 1996).
A few popular scientific names, both comical and sad for those who have any idea of taxonomy, are most often found in articles of this kind. The first name is Dugesiella, most commonly referred to as Dugesiella hentzi. The genus Dugesiella disappeared from the Aphonopelma family a long time ago, and even if it was once assigned to Aphonopelma hentzi (Girard), this cannot be accepted as a credible identification. If a physiologist refers to D. hentzi or A. hentzi, it only means that someone was researching a species of Aphonopelma that someone else thought was a species from Texas.
It's sad, but the name is still walking among physiologists Eurypelmacalifornicum. Genus Eurypelmawas dissolved in another genus some time ago, and the speciesAphonopelmacalifornicumwas declared invalid. These spiders, perhaps, should be attributed toAphonopelmaeutylenum. When you hear these names, it only means that someone thinks that these species are native to California.
Some "scientific" names are really baffling. In the 1970s, someone did a study on a species calledEurypelmahelluo. Apparently, they made a mistake in attributing the species to wolf spiders.Lycosahelluo(now Hognahelluo(Valkenaer)) and changed the name of the genus to make it more similar to the tarantula. God knows who these people investigated.
With varying success, but still physiologists have studied spiders, sometimes even tarantulas, and they have achieved some noteworthy results.
In test tarantulas, it was found that the first (anterior) pair of book lungs controls the flow of blood from the prosoma (cephalothorax), while the second pair of lungs controls blood from the abdomen, before it returns to the heart.
In insects, the heart is predominantly a simple tube that sucks blood from the abdomen, pushes it through the aorta, and ejects it into the head compartment of the insect's body. With spiders, the situation is different. After the blood has passed through the aorta, then through the isthmus between the cephalothorax and abdomen and into the cephalothorax, its flow is divided into what can be defined as a closed system of arteries. It branches and goes to separate parts of the head and legs. Other arteries, called lateral abdominal arteries, originate from the heart on both sides and branch inside the abdomen. From the back of the heart to the arachnoid appendages stretches the so-called. abdominal artery.
When the tarantula's heart contracts (systole), blood is pushed not only forward through the aorta into the cephalothorax, but also from the sides through the lateral arteries and from behind, down through the abdominal artery. Such a system is operable at various levels of blood pressure for the cephalothorax and abdomen. In conditions of increased activity, the blood pressure in the cephalothorax significantly exceeds the blood pressure in the abdomen. In this case, a point is quickly reached when the pressure of the hemolymph in the cephalothorax becomes so great that the blood cannot be pushed from the abdomen into the cephalothorax through the aorta. When this happens, after a certain time, the spider suddenly stops.
Many of us have observed similar behavior in our pets. When the tarantula has an opportunity to escape, some of them immediately fly out of captivity like a bullet. If the tarantula does not reach a place where it feels safe quickly enough, it can run for a while and suddenly freeze, which allows the keeper to catch the fugitive. Most likely, it stops as a result of the fact that the blood stops flowing into the cephalothorax.
From a physiological point of view, there are two main reasons why spiders freeze. The muscles so actively involved in the escape attempt are attached to the cephalothorax. This gives reason to many to believe that the muscles simply run out of oxygen, and they stop working. Perhaps it is. And yet: why does this not lead to stammering, twitching, or other manifestations of muscle weakness? However, this is not observed. The main consumer of oxygen in the cephalothorax of tarantulas is the brain. Could it be that the muscles can work a little longer, but the spider's brain takes oxygen a drop earlier? A simple explanation could be that these maniacal escapees are simply passing out.
The general circulatory system of a spider. When the heart contracts, blood moves not only forward through the aorta and through the pedicele into the cephalothorax, but also laterally through the abdominal arteries down and through the posterior artery behind the heart towards the arachnoid appendages (According to Felix, 1996)
The arachnid class unites over 36,000 species of terrestrial chelicerae belonging to more than 10 orders.
Arachnida- higher chelicerate arthropods with 6 pairs of cephalothoracic limbs. They breathe through the lungs or trachea and, in addition to the coxal glands, have an excretory apparatus in the form of Malpighian vessels lying in the abdomen.
Structure and physiology. external morphology. The body of arachnids most often consists of a cephalothorax and abdomen. The acron and 7 segments are involved in the formation of the cephalothorax (the 7th segment is underdeveloped). In Solpugs and some other lower forms, only the segments of the 4 front pairs of limbs are soldered together, while the posterior 2 segments of the cephalothorax are free, followed by clearly demarcated segments of the abdomen. Thus, the salpugs have: the anterior part of the body, corresponding to the head of trilobites in segmental composition (acron + 4 segments), the so-called propeltidia; two free thoracic segments with legs and a segmented abdomen. Salpugs, therefore, belong to the arachnids with the most richly dissected body.
The next most dismembered detachment is scorpions, in which the cephalothorax is fused, but it is followed by a long 12-segment, like in Gigantostraca, the abdomen, subdivided into a wider anterior abdomen (of 7 segments) and a narrow posterior abdomen (of 5 segments). The body ends in a telson carrying a twisted poisonous needle. This is the same character of segmentation (only without dividing the abdomen into two sections) in representatives of the orders of flagellates, pseudo-scorpions, haymakers, in some ticks and in primitive arthropod spiders.
The next stage of fusion of the trunk segments is found by most spiders and some mites. They have not only the cephalothorax, but also the abdomen, which are continuous undivided parts of the body, but the spiders have a short and narrow stalk between them, formed by the 7th segment of the body. The maximum degree of fusion of body segments is observed in a number of representatives of the order of ticks, in which the whole body is whole, without borders between segments and without constrictions.
As already mentioned, the cephalothorax carries 6 pairs of limbs. The two front pairs are involved in the capture and crushing of food - these are chelicerae and pedipalps. Chelicerae are located in front of the mouth, most often in arachnids they are in the form of short claws (salpugs, scorpions, false scorpions, haymakers, some ticks, etc.). They usually consist of three segments, the terminal segment plays the role of a movable claw finger. More rarely, chelicerae end in a movable claw-like segment or have the appearance of two-segmented appendages with a pointed and serrated edge, with which ticks pierce the integument of animals.
The limbs of the second pair, pedipalps, consist of several segments. With the help of a chewing outgrowth on the main segment of the pedipalp, food is crushed and kneaded, while the other segments make up the genus of tentacles. In representatives of some orders (scorpions, false scorpions), pedipalps are turned into powerful long claws, in others they look like walking legs. The remaining 4 pairs of cephalothoracic limbs consist of 6-7 segments and play the role of walking legs. They end in claws.
In adult arachnids, the abdomen is devoid of typical limbs, although they undoubtedly descended from ancestors with well-developed legs on the anterior abdominal segments. In the embryos of many arachnids (scorpions, spiders), the rudiments of legs are laid on the abdomen, which only subsequently undergo regression. However, in the adult state, the abdominal legs are sometimes preserved, but in a modified form. So, in scorpions on the first segment of the abdomen there is a pair of genital opercula, under which the genital opening opens, on the second - a pair of comb organs, which are equipped with numerous nerve endings and play the role of tactile appendages. Both those and others represent modified limbs. The nature of the lung sacs located on the segments of the abdomen in scorpions, some spiders and pseudoscorpions is the same.
Spider web warts also originate from the limbs. On the lower surface of the abdomen in front of the powder, they have 2-3 pairs of tubercles, seated with hairs and carrying tube-like ducts of numerous arachnoid glands. The homology of these arachnoid warts to the abdominal limbs is proved not only by their embryonic development, but also by their structure in some tropical spiders, in which the warts are especially strongly developed, consist of several segments and even resemble legs in appearance.
Integuments of chelicerae They consist of the cuticle and the underlying layers: the hypodermal epithelium (hypoderm) and the basement membrane. The cuticle itself is a complex three-layer formation. Outside, there is a lipoprotein layer, which reliably protects the body from moisture loss during evaporation. This allowed the chelicerae to become a real land group and populate the most arid regions of the globe. The strength of the cuticle is given by proteins, tanned with phenols and encrusting chitin.
Derivatives of the skin epithelium are some glandular formations, including poisonous and spider glands. The first are characteristic of spiders, flagellates and scorpions; the second - to spiders, false scorpions and some ticks.
Digestive system in representatives of different orders of chelicerates varies greatly. The foregut usually forms an extension - a pharynx equipped with strong muscles, which serves as a pump that draws in semi-liquid food, since arachnids do not take solid food in pieces. A pair of small "salivary glands" open into the foregut. In spiders, the secretion of these glands and the liver is able to break down proteins vigorously. It is introduced into the body of the killed prey and brings its contents into a state of liquid slurry, which is then absorbed by the spider. This is where the so-called extraintestinal digestion takes place.
In most arachnids, the midgut forms long lateral protrusions that increase the capacity and absorptive surface of the intestine. So, in spiders, 5 pairs of blind glandular sacs go from the cephalothoracic part of the middle intestine to the bases of the limbs; similar protrusions are found in ticks, harvestmen and other arachnids. In the abdominal part of the middle intestine, the ducts of the paired digestive gland - the liver - open; it secretes digestive enzymes and serves to absorb nutrients. Intracellular digestion takes place in the liver cells.
excretory system arachnids compared to horseshoe crabs has a completely different character. At the border between the middle and hindgut, a pair of mostly branching Malpighian vessels opens into the alimentary canal. Unlike Tracheata they are of endodermal origin, that is, they are formed at the expense of the midgut. Both in the cells and in the lumen of the Malpighian vessels there are numerous grains of guanine, the main excretory product of arachnids. Guanine, like uric acid excreted by insects, has low solubility and is removed from the body in the form of crystals. At the same time, moisture loss is minimal, which is important for animals that have switched to life on land.
In addition to the Malpighian vessels, arachnids also have typical coxal glands - paired sac-like formations of mesodermal nature, lying in two (rarely in one) segments of the cephalothorax. They are well developed in embryos and at a young age, but in adult animals they more or less atrophy. Fully formed coxal glands consist of a terminal epithelial sac, a loop-like convoluted canal, and a more direct excretory duct with a bladder and external opening. The terminal sac corresponds to the ciliary funnel of the coelomoduct, the opening of which is closed by the remainder of the coelomic epithelium. The coxal glands open at the base of the 3rd or 5th pair of limbs.
Nervous systemArachnida varied. Being connected in origin with the ventral nerve chain of annelids, in arachnids it shows a pronounced tendency to concentration.
The brain has a complex structure. It consists of two sections: the anterior, which innervates the eyes, is the protocerebrum and the posterior is the tritocerebrum, which sends nerves to the first pair of limbs - the chelicerae. The intermediate part of the brain, the deutocerebrum, characteristic of other arthropods (crustaceans, insects), is absent in arachnids. This is due to the disappearance in them, like in the rest of the chelicerae, of the appendages of the acron - antennules, or antennae, which are innervated precisely from the deutocerebrum.
The metamerism of the ventral nerve cord is preserved most clearly in scorpions. In addition to the brain and circumpharyngeal connectives, they have a large ganglionic mass in the cephalothorax on the ventral side, giving nerves to the 2nd-6th pairs of limbs and 7 ganglia, throughout the abdominal part of the nerve chain. In salpugs, in addition to the complex cephalothoracic ganglion, one more node remains on the nerve chain, and in spiders, the entire chain has already merged into the cephalothoracic ganglion.
Finally, in harvestmen and ticks there is not even a clear distinction between the brain and the cephalothoracic ganglion, so that the nervous system forms a continuous ganglionic ring around the esophagus.
sense organsArachnida varied. Mechanical, tactile stimuli, which are very important for arachnids, are perceived by differently arranged sensory hairs, which are especially numerous on the pedipalps. Special hairs - trichobothria, located on the pedipalps, legs and surface of the body, register air vibrations. The so-called lyre-shaped organs, which are small gaps in the cuticle, to the membranous bottom of which sensitive processes of nerve cells fit, are organs of chemical sense and serve for smell. The organs of vision are represented by simple eyes, which most arachnids have. They are located on the dorsal surface of the cephalothorax and usually there are several of them: 12, 8, 6, less often 2. Scorpions, for example, have a pair of median larger eyes and 2-5 pairs of lateral ones. Spiders most often have 8 eyes, usually arranged in two arcs, with the middle eyes of the anterior arc being larger than the others.
Scorpions recognize their own kind only at a distance of 2-3 cm, and some spiders - for 20-30 cm. In jumping spiders (family. Salticidae) vision plays a particularly important role: if males cover their eyes with opaque asphalt varnish, then they cease to distinguish between females and produce the “love dance” characteristic of the mating period.
Respiratory system Arachnids are varied. Some have lung sacs, others have tracheae, and others have both at the same time.
Only lung sacs are found in scorpions, flagellates, and primitive spiders. In scorpions, on the abdominal surface of the 3rd-6th segments of the anterior abdomen, there are 4 pairs of narrow slits - spiracles that lead to the lung sacs. Numerous leaf-like folds parallel to each other protrude into the cavity of the sac, between which narrow slit-like spaces remain, air penetrates into the latter through the respiratory gap, and hemolymph circulates in the lung leaflets. The flagellated and lower spiders have only two pairs of lung sacs.
In most other arachnids (solpugs, haymakers, false scorpions, some ticks), the respiratory organs are represented by tracheae. There are paired respiratory openings, or stigmata, on the 1st or 2nd segments of the abdomen (on the 1st thoracic segment in the salpugs). From each stigma, a bundle of long, thin air tubes of ectodermal origin, blindly closed at the ends, extends into the body (they form as deep protrusions of the outer epithelium). In false scorpions and ticks, these tubes, or tracheas, are simple and do not branch; in haymakers, they form side branches.
Finally, in the order of spiders, both types of respiratory organs are found together. The lower spiders, as already noted, have only lungs; among 2 pairs they are located on the lower side of the abdomen. The rest of the spiders retain only one anterior pair of lungs, and behind the latter there is a pair of tracheal bundles that open outwards with two stigmas. Finally, in one family of spiders ( Caponiidae) there are no lungs at all, and the only respiratory organs are 2 pairs of tracheas.
The lungs and trachea of arachnids arose independently of each other. The lung sacs are undoubtedly more ancient organs. It is believed that the development of the lungs in the process of evolution was associated with a modification of the ventral gill limbs, which the aquatic ancestors of arachnids possessed and which were similar to the gill-bearing ventral legs of horseshoe crabs. Each of these limbs retracted into the body. This created a cavity for the lung leaflets. The lateral edges of the stalk adhered to the body almost along its entire length, except for the area where the respiratory gap was preserved. The abdominal wall of the lung sac, therefore, corresponds to the former limb itself, the anterior section of this wall corresponds to the base of the leg, and the lung leaflets originated from the gill plates located on the back of the abdominal legs of the ancestors. This interpretation is confirmed by the development of lung sacs. The first folded rudiments of the lung plates appear on the posterior wall of the corresponding rudimentary legs before the limb deepens and turns into the lower wall of the lung.
The tracheae arose independently of them and later as organs more adapted to air breathing.
Some small arachnids, including some mites, have no respiratory organs, and breathing takes place through thin covers.

Circulatory system. In forms with clearly expressed metamerism (scorpions), the heart is a long tube lying in the anterior abdomen above the intestines and equipped with 7 pairs of slit-like awns on the sides. In other arachnids, the structure of the heart is more or less simplified: for example, in spiders it is somewhat shortened and carries only 3-4 pairs of ostia, while in haymakers, the number of the latter is reduced to 2-1 pairs. Finally, in ticks, the heart at best turns into a short pouch with one pair of awns. In most ticks, due to their small size, the heart completely disappears.
From the anterior and posterior ends of the heart (scorpions) or only from the anterior (spiders) departs through the vessel - the anterior and posterior aorta. In addition, in a number of forms, a pair of lateral arteries departs from each chamber of the heart. The terminal branches of the arteries pour out the hemolymph into the system of lacunae, that is, into the spaces between the internal organs, from where it enters the pericardial part of the body cavity, and then through the ostia into the heart. The hemolymph of arachnids contains a respiratory pigment, hemocyanin.
Sexual system. Arachnids have separate sexes. The gonads lie in the abdomen and in the most primitive cases are paired. Very often, however, there is a partial fusion of the right and left gonads. Sometimes, in one sex, the gonads are still paired, while in the other, the fusion has already occurred. So, male scorpions have two testes (each of two tubes connected by jumpers), and females have one whole ovary, consisting of three longitudinal tubes connected by transverse adhesions. In spiders, in some cases, the gonads remain separate in both sexes, while in others, in the female, the posterior ends of the ovaries grow together, and a whole gonad is obtained. Paired genital ducts always depart from the gonads, which merge together at the anterior end of the abdomen and open outward through the genital opening, the latter in all arachnids lies on the first segment of the abdomen. Males have various additional glands, females often develop spermatheca.
Development. Instead of external fertilization, which was characteristic of the distant aquatic ancestors of arachnids, they developed internal fertilization, accompanied in primitive cases by spermatophoric insemination or, in more advanced forms, by copulation. The spermatophore is a sac secreted by the male, which contains a portion of seminal fluid, thus protected from drying out during exposure to air. In false scorpions and in many ticks, the male leaves the spermatophore on the ground, and the female captures it with the external genitalia. At the same time, both individuals perform a "nuptial dance" consisting of characteristic postures and movements. The males of many arachnids carry the spermatophore into the female genital opening with the help of chelicerae. Finally, some forms have copulatory organs, but no spermatophores. In some cases, parts of the body that are not directly connected with the reproductive system serve for copulation, for example, the modified end segments of the pedipalps in male spiders.
Most arachnids lay eggs. However, many scorpions, false scorpions, and some ticks have live births. Eggs are mostly large, rich in yolk.
In arachnids, various types of cleavage occur, but in most cases surface cleavage occurs. Later, due to the differentiation of the blastoderm, the germinal streak is formed. Its surface layer is formed by the ectoderm, the deeper layers are the mesoderm, and the deepest layer adjacent to the yolk is the endoderm. The rest of the embryo is dressed only in ectoderm. The formation of the body of the embryo occurs mainly due to the embryonic streak.
In further development, it should be noted that segmentation is more pronounced in embryos, and the body consists of a larger number of segments than in adult animals. So, in the embryos of spiders, the abdomen consists of 12 segments, similar to adult scorpions and scorpions, and there are rudiments of legs on 4-5 anterior segments. With further development, all abdominal segments merge, forming a whole abdomen. In scorpions, the limbs are laid on 6 segments of the anterior abdomen. The anterior pair of them gives genital caps, the second - comb organs, and the development of other pairs is associated with the formation of lungs. All this indicates that the class Arachnida descended from ancestors with rich segmentation and with limbs developed not only on the cephalothorax, but also on the abdomen (prone belly). Almost all arachnids have direct development, but mites have metamorphosis.
Literature: A. Dogel. Zoology of invertebrates. Edition 7, revised and enlarged. Moscow "High School", 1981